愚公产品首登顶刊Cell,助力Wnt信号小体结构功能解析
Cell 顶刊
2026年5月27日,国际顶级学术期刊《Cell》在线发表了上海科技大学许文清教授团队的突破性成果。他们利用冷冻电镜(cryo-EM)技术,首次揭开了Wnt3a信号小体复合物的高清真容,破解了困扰生物学界多年的难题。
 论文链接:https://doi.org/10.1016/j.cell.2026.05.006
论文链接:https://doi.org/10.1016/j.cell.2026.05.006
什么是Wnt信号通路
Wnt信号通路就像是细胞王国的“通信网络”和“指令中心”,调控胚胎发育、成体组织稳态以及干细胞功能等诸多生命过程。典型的Wnt通路包含三个主要成员:
-
信使(Wnt蛋白):细胞外分泌的一类信号蛋白,负责传递指令 。
-
门铃(卷曲蛋白受体Fzd和低密度脂蛋白受体相关蛋白LRP5/6):细胞膜表面的接收器。
-
传话兵(β-catenin连环蛋白):细胞内部的核心效应分子 。
当“信使”Wnt蛋白游动到细胞表面,同时按响“门铃”(结合Fzd和LRP5/6)时 ,细胞内部的“传话兵”β-catenin就会大量积聚,并一路小跑冲进细胞核(指挥部),下达启动特定基因表达的命令(图1)。
这个通路控制着胚胎发育、器官生长以及干细胞的维持 。如果这个“指令中心”传错了信号(比如异常持续激活),细胞就会疯狂失控地生长,最终导致多种疾病的发生 。
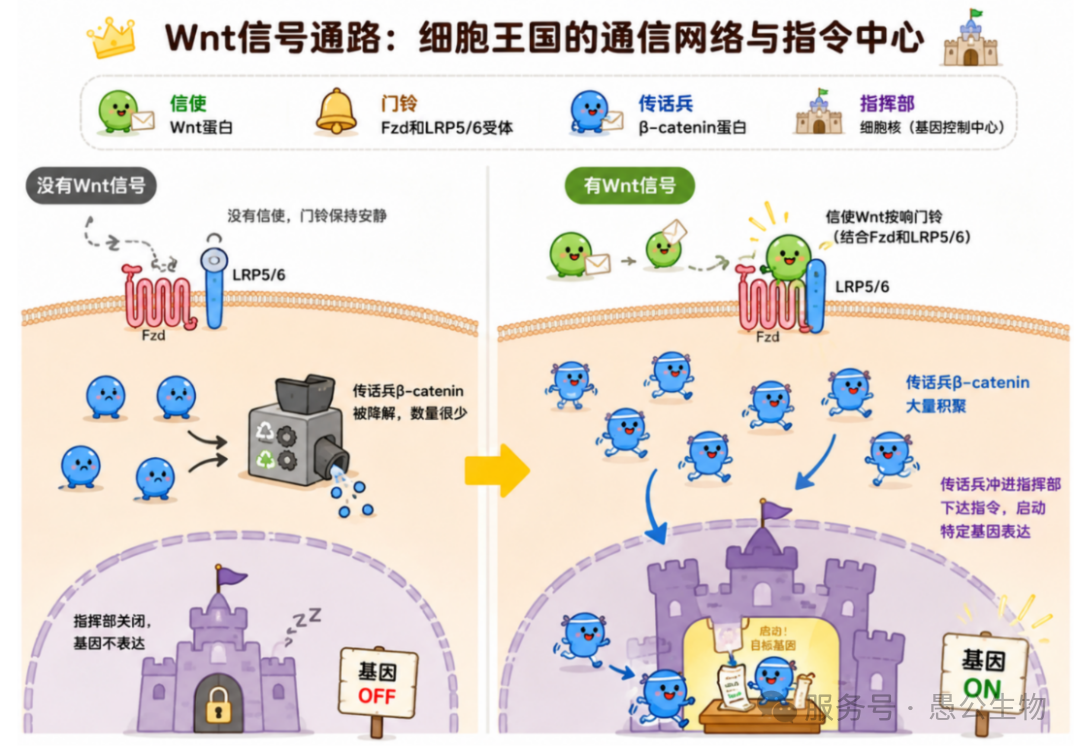
图1 Wnt信号通路示意图
虽然科学家们早在几十年前就知道了这个流程,但由于这些蛋白复合物在体外极不稳定,于是一直有以下几个谜题困扰着科学家们:
✅Wnt3a、Fzd、LRP6三者是如何精准组装在一起的?
✅Wnt蛋白仅仅是“结合”受体就够了吗?有没有隐藏的关键步骤?
✅为什么有些蛋白能阻断这条通路,又该如何人工激活它用于疾病治疗?
六大核心发现
1. 不是“三角恋”,而是“2:4:2”的群舞
以前大家可能简单地认为,一个Wnt蛋白拉着一个Fzd分子和一个LRP分子,就组成了信号复合物。但这项研究通过3套高分辨率的冷冻电镜结构告诉我们,大自然的设计远比这精妙得多。
这个巨型蛋白复合物遵循固定配比:2个Wnt3a蛋白+4个Fzd受体+2个LRP6受体。整个结构的核心骨架是两个Wnt3a形成的同源二聚体,而非单个Wnt蛋白发挥作用(图2)。
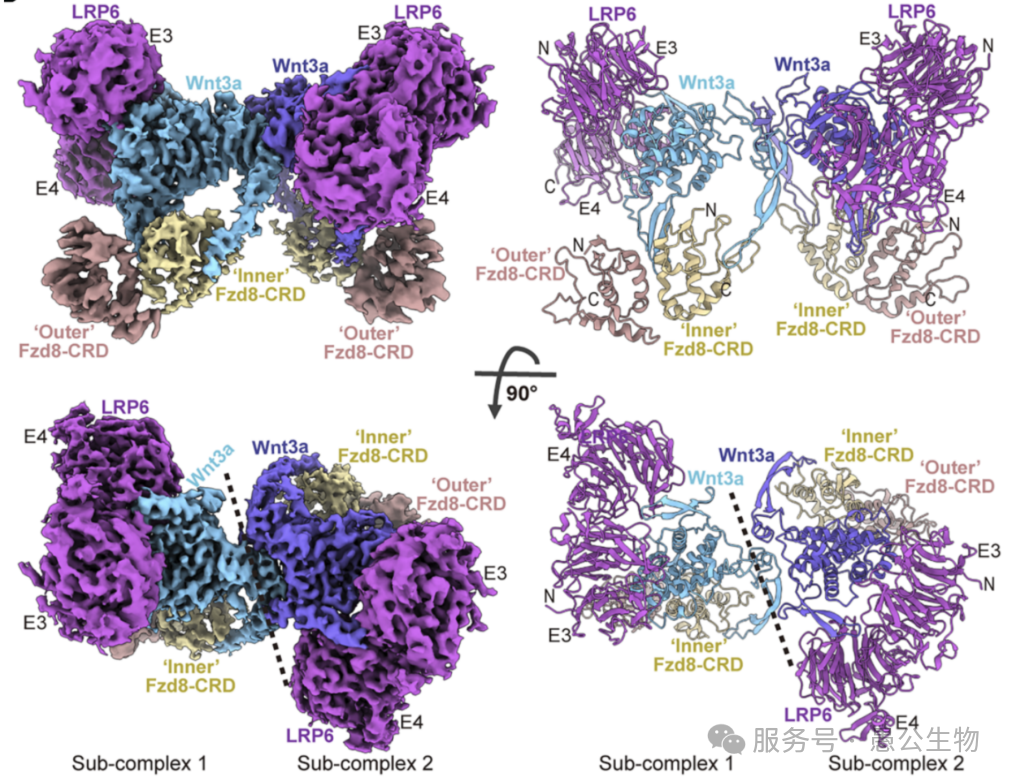
图2 Wnt3a/Fzd8-CRD/LRP6-E3E4 三元复合物的整体结构
更有趣的是结合规则:每一个Wnt3a单体,都会同时结合1个LRP6和2个Fzd受体。大量受体被串联起来,在细胞膜表面形成聚集,这是通路激活的第一步。
2. Wnt3a与LRP6的三大结合界面
Wnt3a和LRP6依靠三处关键接触面牢牢结合(图3A),每一处都不可或缺:
-
结合界面1:依靠疏水作用、盐桥和氢键结合,是两者结合的核心区域(图3B)。
-
结合界面2:通过离子键进一步加固复合物结构(图3C)。
-
特异性结合界面3:LRP6上E3与E4结构域之间的凹槽,只认Wnt3a/Wnt3,其他Wnt蛋白无法结合(图3D )。
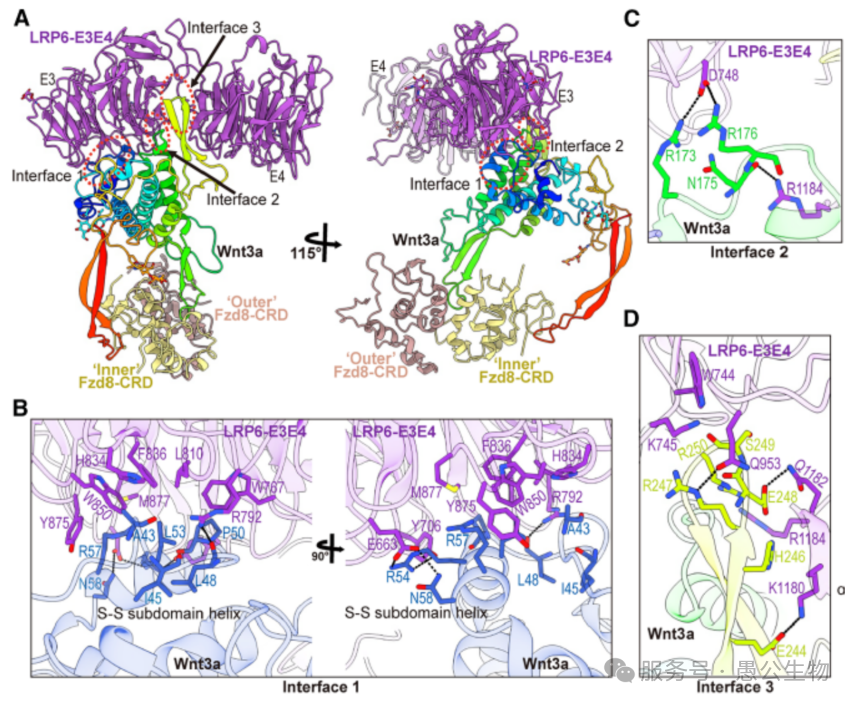
图3 Wnt3a与LRP6结合界面的结构细节
3.找到了“超级活跃版”Wnt3a突变体
Wnt3和LRP6结合界面1上第57位和第58位的氨基酸突变能显著增强信号通路活性,尤其是R57L和R57M两个突变体,可以使Wnt3a的活性提升约8倍(图4)。
这类高活性突变体堪称“天然激动剂”,在干细胞培养、皮肤/骨骼/内脏组织损伤修复、人体类器官培育等再生医学领域,有着巨大的应用潜力。
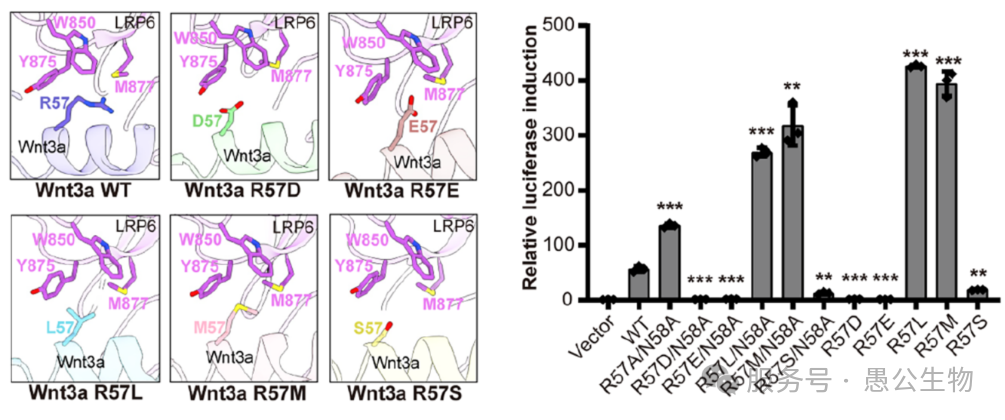
图4 Wnt3a与LRP6结合界面R57突变结构模型与信号活性
4.Wnt3a二聚化是通路激活的“命门”
通过多种生化和细胞实验,证明Wnt3a必须先形成二聚体,才能诱导Fzd、LRP6受体在细胞膜上聚集(图5)。一旦破坏Wnt3a二聚体的结合界面,哪怕蛋白还能结合单个受体,受体也无法聚集,整条Wnt通路会直接罢工(图6)。推翻了以往仅靠配体-受体一对一结合即可激活信号的传统认知。
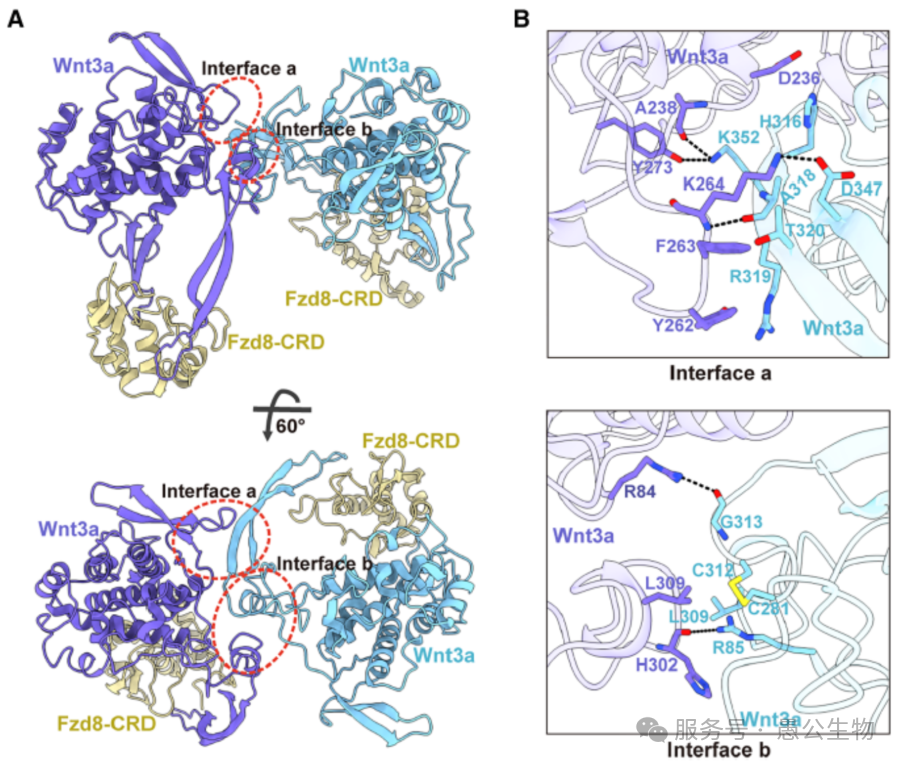
图5 Wnt3a二聚体的结构细节
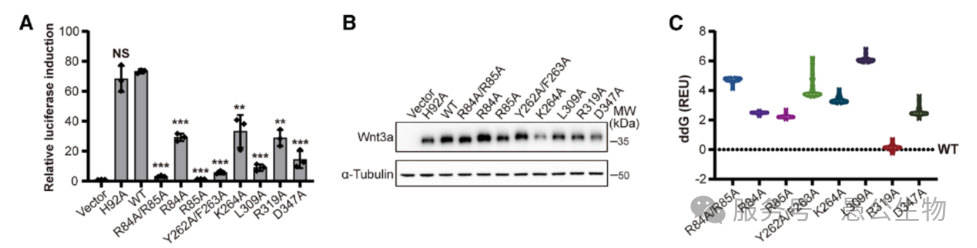 图6 Wnt3a二聚化在Wnt信号小体组装和下游信号传导中起重要作用
图6 Wnt3a二聚化在Wnt信号小体组装和下游信号传导中起重要作用
5.Fzd四聚化:打通胞内外信号的“桥梁”
单个Fzd受体和胞内信号蛋白结合能力很弱,信号根本传不进去。而每个Wnt3a单体结合2个Fzd8-CRD(内侧+外侧),整个复合物中Wnt3a二聚体一次性结合4个Fzd8-CRD形成四聚体。这个四聚体产生叠加效应,高效招募胞内的Dvl信号蛋白,顺利把胞外信号传递到细胞内部,完成跨膜信号传导(图7)。而且这种结合方式在绝大多数Wnt-Fzd识别过程中通用,是Wnt家族的共性机制。
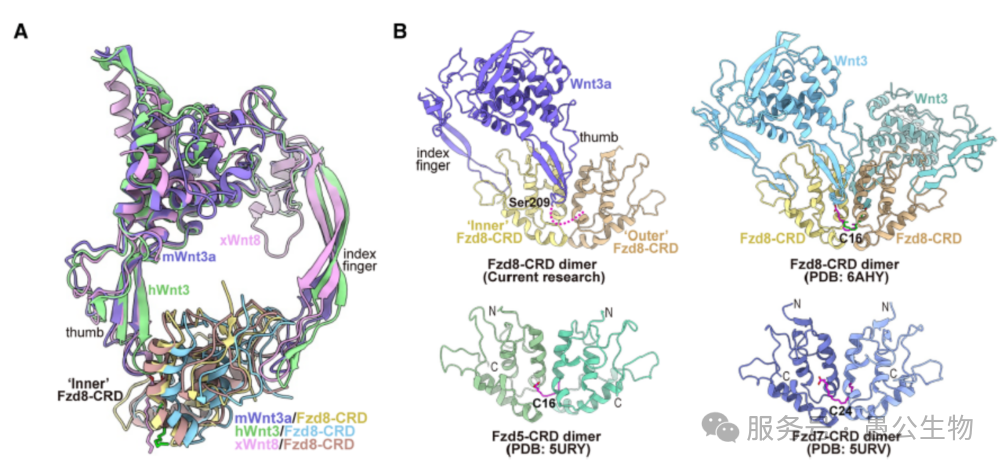
图7 每个Wnt3a蛋白结合2个Fzd-CRD分子
6.解读类似通路的调控密码
将Wnt3a/Fzd8/LRP6复合物与多种此前已报道的相关蛋白结构对比,能够解释很多长期未解的调控机制。
(1) 不同Wnt亚型结合LRP6的偏好性(图8A)
Wnt3a结合LRP6的E3E4结构域,而Wnt8结合LRP6的E1E2结构域,二者结合取向相差近90°;Wnt8缺失Wnt3a关键的S-S亚结构域,是结合区域差异的分子根源,也造就了不同Wnt亚型的不同分工。
(2) DKK1拮抗Wnt3/3a信号的机制(图8B)
Wnt拮抗剂DKK1c与Wnt3a竞争结合LRP6上的同一个结合位点,通过空间占位阻断Wnt3a结合与信号激活。
(3) Wnt-LRP6复合物无法随Wls蛋白分泌的原因(图8C)
Wnt转运蛋白Wls与Wnt3a结合后,LRP6的E4结构域会与内质网膜产生严重空间位阻,因此LRP6不能参与Wnt的内质网分泌过程。
(4) CACHD1结合受体但不激活信号的机制(图8D)
CACHD1也可同时结合Fzd与LRP6,但无法激活Wnt通路。原因有三:①CACHD1仅结合1个Fzd,无法诱导Fzd四聚化;②相比Wnt3a复合物,CACHD1将Fzd与LRP6的间距拉大了25Å以上;③CACHD1结构偏刚性,会将Fzd构象固定为非活性状态。
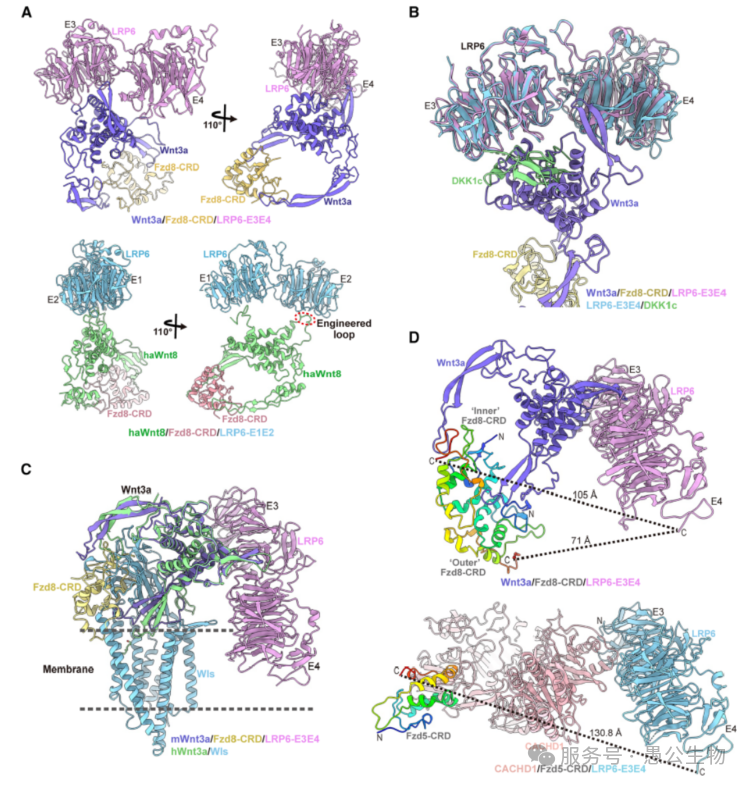 图8 Wnt3a/Fzd8/LRP6复合物结构与相关结构的对比
图8 Wnt3a/Fzd8/LRP6复合物结构与相关结构的对比
Wnt信号小体组装与通路激活的完整分子模型
综上,团队提出了胞外-胞内协同的Wnt通路多层级聚集激活模型(图9):
1. 静息状态(Wnt off)
细胞膜无Wnt配体,胞内β-连环蛋白降解复合物(APC、GSK3B、CK1、Axin)持续降解β-连环蛋白,通路关闭。
2. 激活状态(Wnt on)
(1) 胞外信号小体组装
Wnt3a形成同源二聚体,以2:4:2比例桥接Fzd8与LRP6,诱导Fzd四聚化、Fzd/LRP6膜受体大规模聚集。
(2) 信号跨膜传导(胞内组分招募)
Fzd四聚体高效招募细胞内寡聚化的散乱蛋白Dvl;Dvl进一步招募Axin等组分,组装成完整的跨膜Wnt信号小体,抑制β-catenin降解复合物的功能。
(3)通路最终激活(核内转录)
β-catenin不再被降解,大量入核并结合TCF转录因子,启动靶基因表达,完成信号传导。
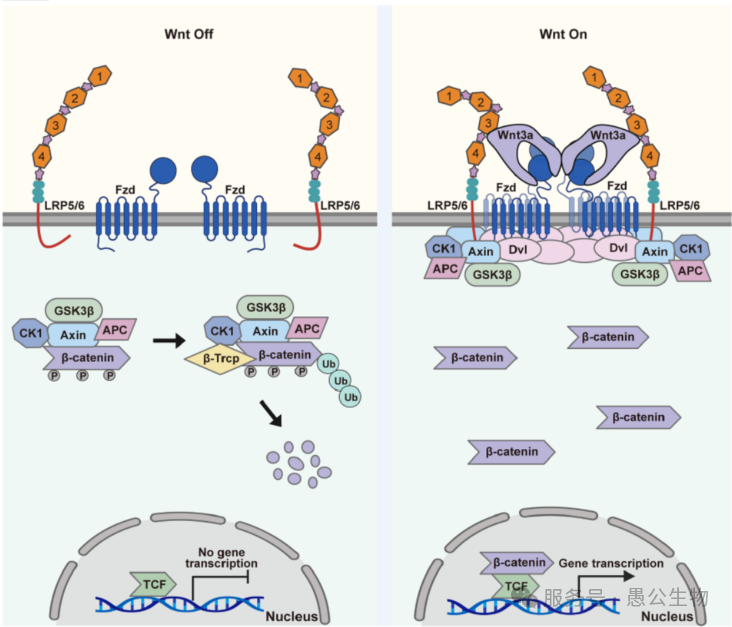 图9 经典Wnt信号通路中Wnt3a信号小体的组装和作用模型
图9 经典Wnt信号通路中Wnt3a信号小体的组装和作用模型
Fzd受体虽然也是G蛋白偶联受体(GPCR)家族的成员,但它在Wnt通路中并不通过经典的构象变构激活,而是依靠配体诱导的受体聚集实现信号激活,这是Wnt-Fzd通路独有的活化模式。
科学意义
理论价值
-
补齐关键短板:第一次解析出天然Wnt3a/Fzd/LRP6完整复合物结构,阐明了Wnt信号小体的组装机制,从结构层面解释了不同Wnt亚型、通路激活剂/抑制剂的作用原理,解答了领域内诸多长期争议的科学问题。
-
更新认知:明确了Wnt二聚化、受体聚集是通路激活的核心,区分了Fzd受体与其他GPCR蛋白的激活机制,为GPCR超家族的分类研究提供新依据。
-
疾病机制研究:为糖尿病、神经退行性疾病、骨骼疾病等Wnt通路相关病症,提供了结构层面的理论支撑。
临床与应用价值
Wnt/Fzd/LRP复合物是生物医药的重要靶点,对于各类类器官的形成与稳态维持也至关重要。本研究为各类临床场景下精准调控Wnt通路提供了理论框架,助力治疗药物、生物制剂、干细胞疗法及组织工程等方向的研发。
幕后功臣
解决复杂载体构建,愚公生物助力科研攻坚
我们在感叹如此精妙的复合物结构之余,不能忽略背后艰巨的分子克隆工作。
Wnt与受体的相互作用亲和力弱、体外表达的复合物很容易异源聚集。为了让体外难稳定的Wnt3a复合物乖乖听话,研究团队设计了精妙的策略:利用雷帕霉素诱导的二聚化系统,将FKBP、FRB结构域分别融合至Fzd8-CRD、LRP6-E3E4末端,中间还加上长达20个氨基酸的柔性Linker,借助雷帕霉素诱导二者靠近,从而稳定三元复合物。后续再突变FKBP关键残基,排除标签蛋白对Wnt3a二聚化与信号活性的干扰,验证天然蛋白的组装特性。随后,为了验证结构和功能,又针对各个结合界面进行了大量密集的定点突变。
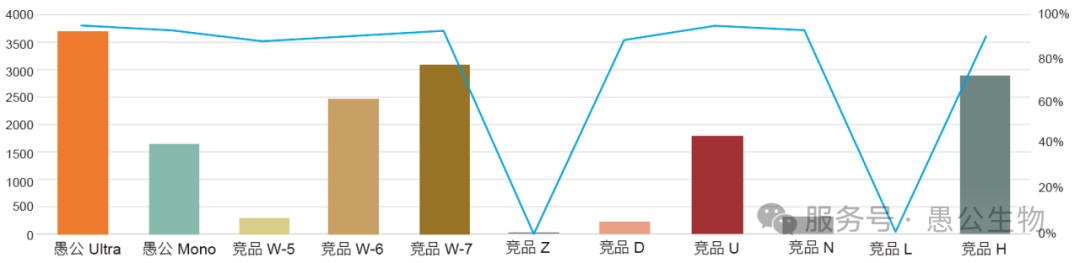
作者选用了愚公生物的LightNing® DNA assembly Mix plus无缝克隆预混液,顺利完成了众多复杂、多片段的质粒构建和定点突变任务。
如今,愚公生物&百时美已经将基于Gibson原理的无缝克隆预混液升级为DNA Assembly Mix Ultra(EG24204)。
相比于上一版本,DNA Assembly Mix Ultra进一步提升了稳定性。在高温、氧化、反复冻融或干冰/ 二氧化碳暴露等不利条件下,依然可以获得足够数量的阳性转化子。
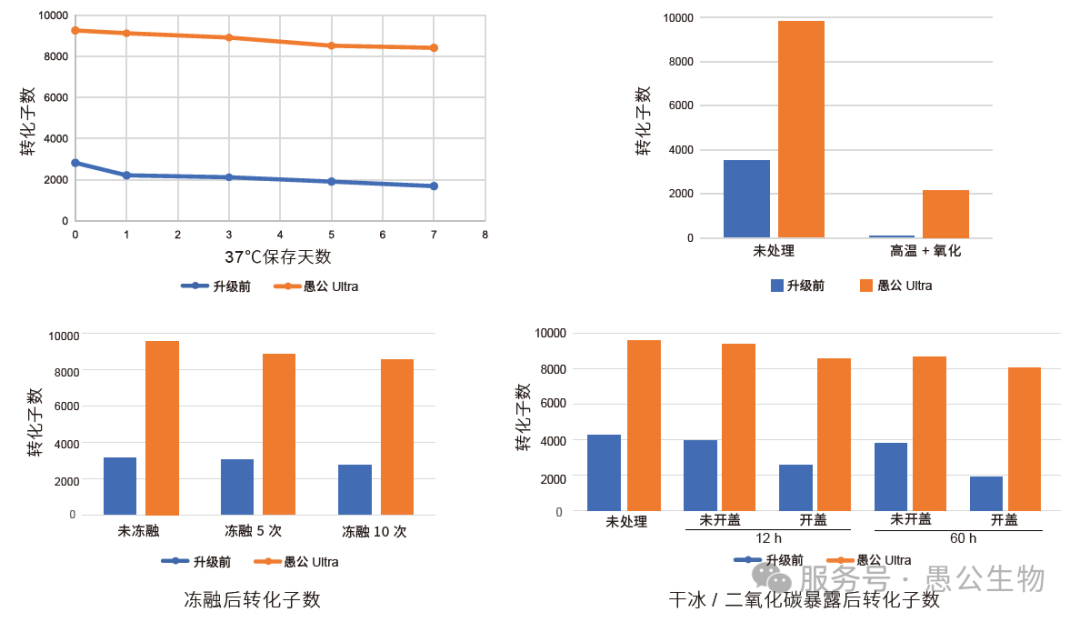
10 片段连接的条件下,升级版DNA Assembly Mix Ultra 也能得到足够的转化子数和很高的阳性率。